�_�o�זE�����͂Ȃ�Neuron�ESynapse�EReceptor
�_�o�זE�̐�
��]�玿�̐_�o�זE�̐��͂��悻100������180�����炢����A
��ʂɂ͕��ϒl���Ƃ��đ�]�玿�̐_�o�זE�̐�140���ł���
�Ƃ���Ă��܂��B
��]�玿�̐_�o�זE���́A�`���p���W�[�Ŗ�80���A�A�J�Q�U��
�i�j�z���U���̒��ԁj�Ŗ�50���Ƃ���Ă��܂��B�@
���ӂ��ׂ��_�́A��]�玿�̐_�o�זE�̐��ł���A�]���邢�͒����_�o�i�]�ƐҐ��j�S�̂̐��ł͂Ȃ��Ƃ������Ƃł��B��]�̓����ɂ͐_�o�זE�̏W�܂����_�o�j������A���]��Ґ��ɂ���R�̐_�o�זE������,���]�����ł�1,000���ȏ�̐_�o�זE������Ƃ����T�Z������܂��B�]���āA�����_�o�S�̂̐_�o�זE�̐���1,000����2,000���̊ԂƐ��肳��܂��B
�g�̂̐����Ƌ��ɐ_�o�זE�̃l�b�g���[�N�������ɐ����B�������Ԃ́A10��㔼�܂łƍl�����Ă��܂��B
�������~�܂�n�߂�̂�4�Έʂ���ŁA���o�A���o�A�{�����o�A�����ʼn^���A����Ɋւ��镔�ʁi�^����E�����j�A���f�E�v����i�镔�ʁi�O���O��j�̏����ł��B
�ŋ߂̌����ŁA�^���A�����A�p�����鎖�ŐV�����_�o�זE�l�b�g���[�N�̊������͉\�Ɖ����Ă��܂����B
�זE��1�N�ʂŐV�����זE�Ɠ���ւ��܂����A�]�A�_�o�זE�A�S���i�S�؍זE�j�͓���ւ��A�Đ��ł��Ȃ��ƌ����Ă��܂��B
Page Contents�@�i�ڍׂ́A�����������N���b�N�j
�@�@�@�E�_�o�זE�i ���� �E ����ˋN�j�@�@�@�E�V�i�v�X�@�@�@�E�_�o�זE�̎���@�@�@�E�O���A�זE�@�@
�@�@�@�E�זE�ϐ��i�E���E�Đ��j�@�@�E�_�o�זE���튯
�@�@�@�E�G���h�\�[���@�@�E�~�g�R���h���A�@�@�E�I�[�g�t�@�W�[�@�@�E���r�L�`���A�v���e�A�\�[���@�@
�@�@�@�E�G���h�T�C�g�|�V�X�@�@�E�G�L�\�T�C�g�|�V�X�@�@
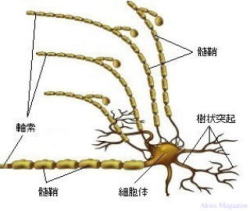
�_�o�זE
�_�o�זE�@�i ���`�B�j���[�����ŁA�����E����ˋN�Ɏ}������j
| �_�o�זE�́A���`�B�ɓ��������זE�ł��B��̐_�o�זE����͒����u�����v�i���������A�_�o�`�B������ �@���ˁj�ƁA���G�Ɏ}�����ꂵ�Ă���u����ˋN�v�i���ザ�傤�Ƃ����A�_�o�`�B�����̎�e�j�ƌĂ�� �@�ˋN���o�Ă��āA�����̓ˋN�͕ʂ̐_�o�זE�ƂȂ��荇���A���G�Ȑ_�o��H�ԁi�l�b�g���[�N�j���`���� �@�Ă��܂��B1�̐_�o�זE�ɂ͖�1���̎���ˋN���t���Ă���ƌ����Ă��܂��B�܂��A�זE���ɕ�܂�j�A �@�~�g�R���h���A�A���\�\�[���Ȃǂ̍זE���튯�������Ă��܂��B �@�����́A���𑗂�o���ˋN��1�{�A�������}�Ɩ��[�͎}������i�I�����}�j�Ă��܂��B �@����ˋN�́A�������ˋN�ōזE�̂��畡���}�����ꂵ�Ă��܂��B �@���̎�悪���Ȃ�����ˋN�͏�������A�c��������ˋN�̎}���������͂̑��������ɑ��₷���ƂŁA �@�������I�Ȑ_�o��H���`������܂��B �@1�̐_�o�זE�͂��ꂼ��1�����̐_�o�זE�ƘA������荇���Ă��܂��B�_�o�זE���ł́A�d�C�̗��ꂪ �@����`���A�_�o�זE�Ɛ_�o�זE�̐ڍ������̓V�i�v�X�Ԍ��ƌĂ��킸���Ȍ��Ԃ�����A���̕����ł� �@�_�o�`�B���������̐_�o�זE�ɏ���`�B���܂��B �@�O���A�זE�́A�_�o�n���\������זE�ł͗L��܂��A�_�o�זE�ɑ��ĉh�{�f�̋�����ʒu�̌Œ���s�� �@�Ă���A���̂̓�����O���̊����q�̕ω��ɂ�����炸���̂̏�Ԃ����ɕۂ����Ƃ��������i�P�퐫�j �@�������Ă��܂��B 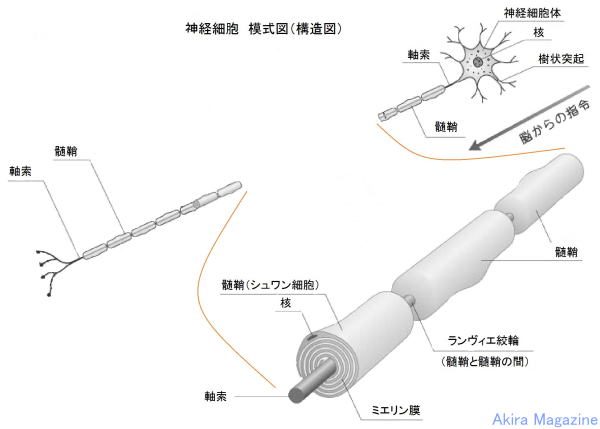 ���≻�i�~�G�������j�A�_�o�`�B��H�̌`���ɕt���āF �@�_�o�זE���玟�̐_�o�זE�ւ̐_�o�`�B�͓d�ʂōs���A�_�o�I�������ŃV�i�v�X���E�ɂ��V�i�v�X�Ԍ��� �@�V�i�v�X���E�̒��̐_�o�`�B���������o�������ˋN�i�_�o�`�B����_�o�זE�j�̎�e�̂��ւĐ_�o�`�B �@���s���鎖���m���Ă���B �@�_�o�זE���V��������鎞���́A20�ΈʂƂ���Ă��Đ_�o�`�B��H�̌`���͏I���A���̌�A�g�p�p�x�� �@���Ȃ��_�o�`�B��H�i�_�o�זE�j�͏��ł���Ƃ���Ă��܂����B �@�ŋ߂̌����ŁA�]���̈ꕔ���ʁi�C�n�j�ł́A�_�o�`�B��H�i�_�o�זE�j�̐V���Ȍ`�����s���邱�ƁA���≻ �@�i�~�G�������j�𑣐i�����鎖�������ɂ������Ă��܂����B �@����Ƃ́A�_�o�זE�̎������މ~����̑w�i�������b�����������≏�́j�Ŏ����A����̌a�����鎖 �@�i�������n�j�ɂ��_�o�`�B����荂����������B �@���≻�i�~�G�������j�́A�����_�o�ł̓O���A�זE�i�I���S�f���h���T�C�g�j�A�����_�o�ł̓V�������זE �@���狟�������������̌a�ɒB����ΐ���`�������i����܂��B |
�V�i�v�X
�_�o�זE�̂Ȃ��ڃV�i�v�X
| �_�o�זE�Ԃ̂Ȃ��ڂɊւ��\�����V�i�v�X�ƌĂ�Ă��܂��B �_�o�זE�̎������d�ʂ��`���A�_�o�`�B��������܂ꂽ�V�i�v�X���E���זE���i�V�i�v�X�O���j�ɐڂ����d�ʈˑ����J���V�E���C�I���`�����l�����J���J���V�E������������V�i�v�X���E���Ǝ����I�����̍זE���Ƃ� �Z�����ăV�i�v�X���E���ɂ���_�o�`�B�������V�i�v�X�Ԍ��ɕ��o����A����ˋN�̎�e�̂Ɏ�荞�܂ꌋ�����鎖�ɂ����`�B�����B �V�i�v�X���E�̑傫����50nm�قǁA�V�i�v�X�Ԍ��i�����j��20nm�قǁB(1nm=10������1m)�A �V�i�v�X�Ԍ��̓`�B���Ԃ�0.1�`0.2�~���b�قǁB ���`�B�@���F �@1�F�_�o�זE�̎�������d�ʂ��`���A�����I�����ɓ��B����B �@2�F�d�ʂɂ�莲���I�����Ɉʒu����d�ʈˑ����J���V�E�� �C�I���`���l�����J���B �@3�F�J���V�E�� �C�I���������I�����ɗ������A�V�i�v�X���E���זE���ɐڂ��Đ_�o�`�B�������זE�O�ɊJ�� �@�@ ���o�B �@4�F�_�o�`�B�����̓V�i�v�X�Ԍ����g�U���A����ˋN�i�j�̍זE����ɕ��z����_�o�`�B������e�̂� �@�@ �����B �@5�F����ˋN���̃V�i�v�X�㖌�̃C�I�� �`���l�����J���A�זE�����O�̓d�ʍ����ω����܂��B 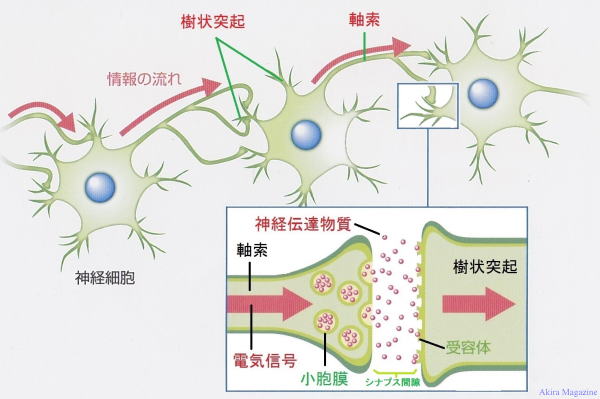 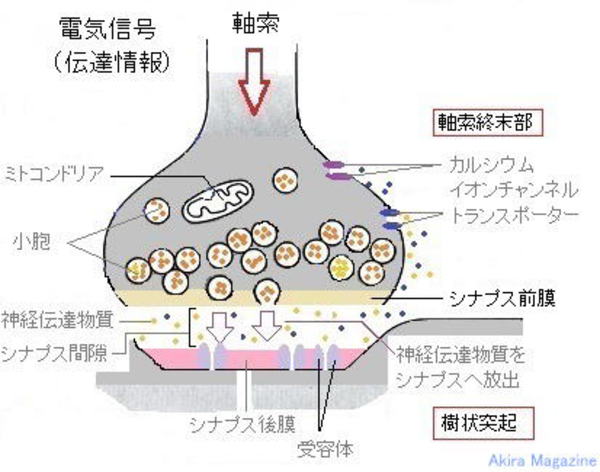 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�_�o�זE �����E�V�i�v�X�E����ˋN�} |
�_�o�זE�̎��
�_�o�זE�̎�ށE����
| �_�o�זE�̓����ނ�������o�A��݁A�^���̂R��ނɕ������܂��B�ȒP�Ɍ����ƁA�O����̎h�������̂����o�_�o�זE�A�h�����ؓ��ɓ`����̂��^���_�o�זE�A�_�o�זE �Ԃł̏��`�B������̂���ݐ_�o�זE�ł��B���̉�ݐ_�o�זE���A�]�Ȃǂ̒����_�o�n���`�����Ă��܂��B ���`�B�͓d�C�M���i�����d�ʁj�ɂ��o�́A���͂ƍs���A�_�o�זE�i�����A����ˋN�j�̖��[�ł́A �V�i�v�X�ƌ������Ԃ����肻�̊Ԃ͐_�o�`�B�����ŏ�`�B����Ă��܂��B ���o�_�o�זE�̎��
|
||||||||||||||||||
�O���A�זE
�O���A�זE�̎�ށE����
| �����_�o�n��Ґ��ɑ��݂��Đ_�o�זE�i���o�E�^���E��݁j�̈ێ��Ɋ֗^����זE�Q�ŁA�_�o�זE�ɑ��A �h�{�f�̋�����_�o�זE�̈ʒu�Œ�ȂǁA�̉��⌌���A�̉t�̐Z������pH�Ȃǂ��͂��ߕa����������E�C���X �Ƃ������ٕ��̔r���A�C���Ȃǐ��̋@�\�S�ʂɋy�ԍP�퐫�̈ێ���S���זE�ŁA�Ɖu�n�̂悤�ȐU�镑��������זE�B�_�o�זE���̖�10�{�A�̐ςł͔]�̔������߂�Ɛ�������Ă���B 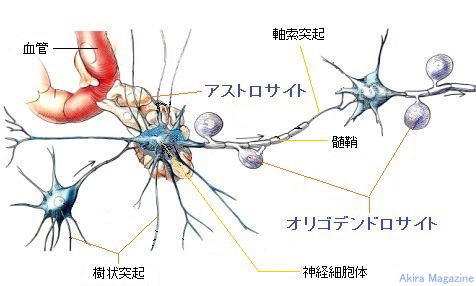 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�����_�o�n�̃O���A�זE�} �O���A�זE�̎��
|
||||||||||||||||||||||||||||
�זE�ϐ�
�_�o�זE�̕ϐ��E�E���E�Đ��@
| �_�o�זE�̈ꕔ����������ƁA���̏ꏊ�����זE�̂��牓�����͕ϐ����ĉ��Ă��܂����s���ϐ��B �זE�̂̂��鑤�ɂ��ϐ����i�s���邱�Ƃ�����A������t�s���ϐ��B�܂��A�_�o�זE�݂͌��ɐ_�o�h�{���q�Ȃǂ�����肵�Ă���A�V�i�v�X�Őڑ����Ă���זE����ꂽ�ꍇ�ɂ��A�_�o�h�{���q�̕s������זE�����N�������Ƃ�����܂��B���̏ꍇ���A�O�V�i�v�X�זE�������Ƃɂ���V�i�v�X�זE�����ʏꍇ�����s���ϐ��A��V�i�v�X�זE�������Ƃɂ��O�V�i�v�X�זE�����ʏꍇ���t�s���ϐ��ƌĂԂ��Ƃ�����܂��B 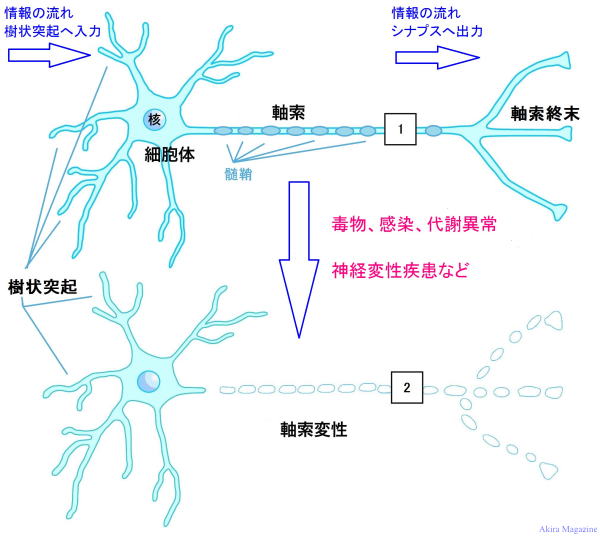 ��Q�̒��x���������čזE�̂�����ł��܂��ƁA���̐_�o�זE�͂��͂�Đ��s�\�B�������A�����_�o�i�p��W�Q�Ɓj�̏ꍇ�A�זE�̂������Ă���A�Ăю�����L���ĖړI�זE�Ƃ̌������ł��邱�Ƃ������A���̉ߒ��ɂ́A��ꖌ��V�������זE�̊֗^���K�v�Ƃ���Ă��܂��B����A�����_�o�ɔ�ׂĒ����_�o�͂قƂ�ǍĐ� �\�͂��Ȃ��A�]��Ґ��̑����͐��U�ɓn���Č��ǂ��c�����Ƃ����Ȃ��Ȃ��B |
�_�o�זE���튯
�_�o�זE���튯 �i �j���́E���E�́E���{�\�[���E�S�W���́E���\�\�[�� �j
| �זE���튯 (Organelle) ���A �^�j���������זE���튯�B�j���́A���E�́A�S���W�́A���\�\�[���A �@�~�g�R���h���A�A�y���I�L�V�\�[�����̖��ň͂܂ꂽ�\���́B �@���E�́A�S���W�́A�G���h�\�[���A���\�\�[���́A���E����čזE���ƘA���������Ă���B �@���̓��O�ŗl�X�ȕ����̔Z�x���R���g���[�����āA�G�l���M�[���Y�i�d�q�`�B�n�j�A�����̒����Ȃǂ��s���� �@����B �@�E�j�����icell nucleus)�ɂ́A��`���ł���DNA�̂ق��A�j�^���p�N���ARNA�i���{�j�_�j���܂܂�� �@�@���܂��B �@�E���{�\�[���iRibosome�A�^���p�N���̐����H��j�́A�זE�j��DNA���`��RNA�i�v�}�Aen: �@�@ MessengerRNA�j��ǂݎ��A�]��RNA�ien: Transfer RNA (TRNA)�Ɍ��т����A�~�m�_���珊��� �@�@�^���p�N���ɑg�ݗ��Ă�B�i�זE�����\������^���p�N���A����^���p�N���������j �@�E���E���iendoplasmic reticulum�j�́A���{�\�[���ō��ꂽ�^���p�N���𗧑̍\���i�܂肽���ށj�� �@�@�ւ���Ă��܂��B �@�E�e�ʏ��E���i���߂傤�ق�����: rough-surfaced endoplasmic reticulum, rER�j�ł́A���{�\�[���� �@�@�t�����Ă���A�S���W�̂�\�\�[���A���E�́A�זE�������\������^���p�N������сA����^���p�N���� �@�@�����B �@�E�S�W�����iGolgi body�A�^���p�N���̍ŏI�����H���j�́A���{�\�[�� �� ���E�� �� �S�W���̂։^��A �@�@����^���p�N����זE�O�^���p�N���̓����C����A���E�́i�e�ʏ��E�́j�ɂ�萶�Y���ꂽ�e��O��� �@�@�^���p�N���̉��w�I�C���i�^���p�N���������j���s���^���p�N���ނ��A���������A���\�\�[�����邢�� �@�@�זE���ɂ��ꂼ��U�蕪���铭�������B �@�E�G���h�\�[���́A�����̑I�ʁA�����ė��p�̋@�\�B�ʍ��ɋL�ځB �@�E�~�g�R���h���A�ɕt���Ă͕ʍ��ɋL�ځB �@�E���\�\�[�� (Lysosome) ���A �̍����q�̕����ƍė��p�B�܂��A�זE���̕��u�Ƃ��Ă����łȂ��A �@�@���ɂ��l�X�ȋ@�\��L���Ă��܂��B���\�\�[���̃G�L�\�T�C�g�\�V�X�i�זE�O�ւ̕����̕��o�j�́A���� �@�@�����זE���̏C���A�זE�O�}�g���b�N�X�̕����ȂǂɊ֗^�B���\�\�[�����̓��ߐ����i�ilysosomal �@�@ membrane permeabilization�j�ƍזE���Ƃ̊֘A������Ă��܂��B�܂��A �@�@ ���\�\�[���͍זE���̉h�{��ԁi�A�~�m�_�Ȃǁj�����m�����Ƃ��Ă��d�v�ŁA����ɍזE���̃A�~�m�_ �@�@�Z�x�����m����Z���T�[�^���p�N���̑������\�\�[������ɋǍ݂��Ă��܂��B�זE���̃A�~�m�_�Z�x���㏸ �@�@����ƁA�זE�����E��ӁE�^���p�N�������Ȃǂ̗l�X�ȍזE�@�\�𐧌䂷��d�v�ȃV�O�i�����q�ŁA �@�@���̊������̓��\�\�[������ŋN����܂��B �@�@���\�\�[���̓��o�͎�X�̉��������y�f���܂݁A�זE���O�����̕����@�\��S���Ă��܂��B �@�@������̓G���h�T�C�g�[�V�X�A�}�N���I�[�g�t�@�W�[�Ȃǂ̌o�H�ɂ���ă��\�\�[���ɗA������܂��B �@�@�������ꂽ���̂̂����L�p�Ȃ��̂́A�זE���ɋz������܂��B�s�p���̓G�L�\�T�C�g�[�V�X �@�@(�זE�O�֕��o�j�ɂ���Ĕp������邩�A�c�]���́iresidual body�j�Ƃ��čזE���ɗ��܂�܂��B �@�@�ꎟ���\�\�[���iprimary lysosome�A�ꎟ�������́j�F �@�@��������܂܂Ȃ����\�\�[�����w���A�������ψ�ȍ��d�q���x�����B �@�@���\�\�[���isecondary lysosome�A�������́j�F �@�@�G���h�\�[���A�t�@�S�\�[���A�I�[�g�t�@�S�\�[���ƗZ������������܂ꎟ���\�\�[���B �@�@���\�\�[���́A����Ɋ�̗A���o�H�ɏ]���ăt�@�S���\�\�[���iphagolysosome�A�H���Z�����́j �@�@�I�[�g���\�\�[���iautolysosome�A���ƐH�Z�����́j�ƌĂ�Ă��܂��B �@�@�זE���ɂ͗l�X�ȃ��\�\�[��������A���̑傫���E�`��͗l�X�ɈقȂ��Ă��܂��B �@�@���\�\�[�����ܗL������������y�f�Q�͎_���������Ō����ǂ����������������Ă���A���\�\�[�������� �@�@���f�C�I���w���̓v���g���|���v�̓����ɂ���� ���f�C�I���w���lpH5 �O��Ǝ_���ɕۂ���Ă��܂��B �@�@ �ipH7������ �j���̂��Ƃɂ���āA������Ԃ̑��̍זE�����ł̓��\�\�[�����܂މ��������y�f�͕s�� �@�@���ƂȂ�A�s�K�v�Ȕ�����h���ł��܂��B �@�@���C�\�\�[���i�����\�[���j�ɏ�Q���N��ƃ~�g�R���h���A�̓S��ӂɉe�����y�ڂ��A������������ �@�@���C�\�\�[���y�f�A�O���R�Z���u���V�^�[�[(GBA)�̌��R�ɂ��S�[�V�F�a���a�B �@�@�S�[�V�F�a�́A�̑����B���̔��A���������ɂɂ��n���ǁA������ς��낭�Ȃ�A�p�[�L���\���j�Y�� �@�@���������₷���S���̒�����������B 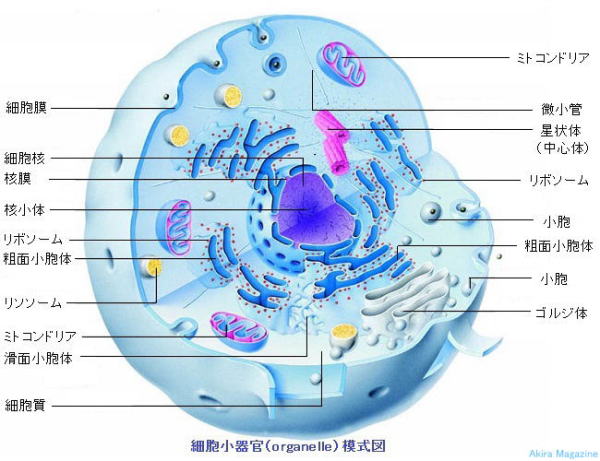 �^���p�N���̍����E�ُ�^���p�N���̏����A�i���Ǘ� �@�^���p�N���̓A�~�m�_�������������܂��B�V���ɍ�낤�Ƃ���^���p�N���́A���̐v�}�iRNA�j�� �@������܂��BRNA�͍זE�j��DNA�ɂ��R�s�[���ꃊ�{�\�[���ō���܂��B���̌�^���p�N���͏��E �@�̂Ő܂��݁i���̍\���j����܂����A�ُ�^���p�N���A�������^���p�N�������ꂽ���ɐ���̃^���p�N�� �@�ɖ߂����߂ɕ��q�V���y��������������Ȑ܂��݂ɖ߂��̂ł����A��ʂɈُ�A�������^���p�N�����o���� �@�ꍇ�i�זE�̓��A���E�̓��j�ɂ́A�ُ�^���p�N���̋ÏW�ɂ��זE���̂����ɂ܂��B���̌��ۂ��t�H�[�� �@�f�B���O�a�i�A���c�n�C�}�[�^�F�m�ǁA�p�[�L���\���a�A�v���I���a���j�ł��B���̎���������邽�߂� �@���E�̃X�g���X�����ƌ����@�\������܂��B�������s���鏇�Ԃ́A �@�@�@ PERK�ɂ��ُ�^���p�N���̍�����~�A �@�@�A ATF6�ɂ��V���y����RNA�����{�\�[���Ɏw�����ď��E�̂ɐ���Ȑ܂��݂��w�߂����B �@�@�B IRE1��XBP1�ɂ��^���p�N���̕����ƍĐ��ɂ��`�����̕i���Ǘ����s���Ă��܂��B �@�i�]�ʈ��q:ATF6�AXBP1�͏��E�̂ɑ��݂��Ĉُ�^���p�N�����o�����ATF6���藣����זE�j�֍s���זE �@�j��ATF6�����m���ăV���y����RNA�����{�\�[���Ɏw�����ď��E�̂ɐ���Ȑ܂��ݎw�����w�߂���܂��j |
�G���h�\�[��
�G���h�\�[���@�@�i �זE���̖������A�� �j
| �G���h�\�[�� (Endosome)�́A�G���h�T�C�g�[�V�X�ɂ���čזE���ւƎ�荞�܂ꂽ�l�X�ȕ����̑I�ʁE���� �@�E�ė��p�Ȃǂ𐧌�B�G���h�\�[���͌`�ԓI�ȓ�����@�\�I�Ȃǂ̓��������ƂɁA �@�@�@�����G���h�\�[���iearly endosome�j �@�@�@����G���h�\�[���ilate endosome�j �@�@���T�C�N�����O�G���h�\�[���irecycling endosome�j�ɑ�ʂ����B �@�@�G���h�\�[���͖��A���̒��p�n�_�Ƃ��ċ@�\���邱�ƂōזE���̕����A�����i�邱�Ƃ���A�_�o�@�\�� �@�@�͂��߂Ƃ���l�X�Ȑ������ۂɂ����ďd�v�Ȗ������ʂ����Ă���B 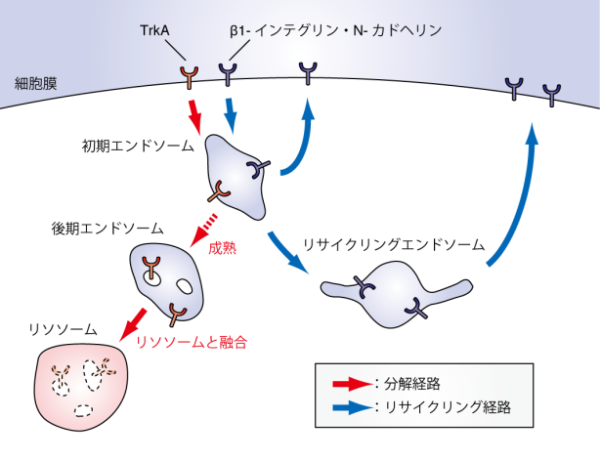 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�G���h�\�[���ɂ��`���������o�H�} �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�o�T�摜�FWikipedia Commons �@�����iTrkA�j�͕����o�H�ցA�זE���ōė��p����镨���i��1-�C���e�O�����AN-�J�h�w�����j�� �@���T�C�N�����O�o�H�ւƑI�ʂ���Ă����B�����G���h�\�[���ɂ����ĕ����o�H�ւƑI�ʂ��ꂽ�����́A �@�����G���h�\�[���ɗ��܂�B �@�����ď����G���h�\�[��������G���h�\�[���ւƐ��n����ƁA����G���h�\�[�������\�\�[���ƗZ�����A �@���o���E���Ɩ��ђʌ^�̃^���p�N�������������B �@�����G���h�\�[���ɂ����ă��T�C�N�����O�o�H�ւƑI�ʂ��ꂽ�����́A�����G���h�\�[�����璼�ځA���邢�� �@�j�ߖT�̃��T�C�N�����O�G���h�\�[���ւƗA�����ꂽ�̂��A�זE���ւƖ߂���ė��p�����B |
�~�g�R���h���A
�~�g�R���h���A�@�@�i �G�l���M�[�̎Y���A�A�|�g�[�V�X�U���Ɋ֗^ �j
| �~�g�R���h���A (mitochondria�j�́A�^�j�����̍זE���튯�̈�B��d�̐��̖�����Ȃ�A�Ǝ���DNA �@�imtDNA�j�������A����A���B����B�~�g�R���h���A�͍זE�̒��Ŏ_�f�i���t�ɂ��^�ꂽ�j�ɂ�蓜�� �@���b��R�����鎖�ɂ��M�̎Y���i�̉��j�ADNA�����A�X�g���X�ɂ��A�|�g�[�V�X�U���A���߁A�J���V�E �@�������A�זE���̃J���V�E���Z�x�̒����Ɋ֗^���Ă���B�i�~�g�R���h���ADNA�imtDNA�j�̓A�f�m�V�� �@�O�����_�iATP�j�ȊO�̃G�l���M�[�Y���Ɋ֗^�jmtDNA�́A��n��`�q�ŕ�e����p����܂��B �@�~�g�R���h���A�̍\���́A�O���Ɠ����̎������ɕ�܂�Ă���A�����̓����̓}�g���b�N�X�ƌĂ�A���� �@���畽��i�N���X�e�j������B�O���ɂ́A���^���p�N���i�|�����j������A���q��5000�ȉ��̕��q������ �@�ł���悤�ȃ`���l�����`�����Ă���B �@�����ɂ́A�_���I�����_�Ɋւ��ċz�������̂Ȃǂ̍y�f�Q���K���I�ɔz�Ă���B�܂��A�}�g���b�N�X�� �@�̃^���p�N���A�����u��~�g�R���h���A�̕���E�Z���Ɋւ��^���p�N���Q�Ȃǂ����݁B �@�}�g���b�N�X�ɂ́ATCA��H�i�N���u�X��H�E�N�G���_��H�j����_���Ȃǃ~�g�R���h���A�̑�Ӌ@�\�Ɋւ� �@��y�f�Q�A���{�\�[���AtRNA�A�]�ʈ��q��|����q�Ȃǂ����݂��Ă���B �@�~�g�R���h���A�̋@�\�́A�d�q�`�B�n�A�����_���ɂ��ATP�̎Y���iADP�̃����_���j�B�זE�̊����ɕK�v�� �@�G�l���M�[�̂قƂ�ǂ͒��ځA�ԐړI�Ƀ~�g�R���h���A����ATP�̌`�ŋ��������B ���̑��̋@�\�Ƃ��Ă� �@�A�X�e���C�h��w���̍����Ȃǂ��܂ޗl�X�ȑ�ӁA�J���V�E����S�̍זE���Z�x�̒��߁A�זE������ �@�A�|�g�[�V�X�̒��߂Ȃǂɂ��傫���ւ���Ă���B 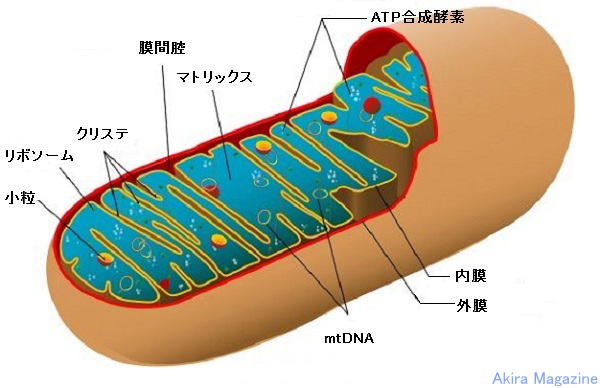 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�~�g�R���h���A�͎��} |
�I�[�g�t�@�W�[
�I�[�g�t�@�W�[�@�i �`���������̈�� �j
| �I�[�g�t�@�W�[ (Autophagy)�́A�זE�������Ă���A�זE���̃^���p�N�������邽�߂̎d�g�݂̈�ŁA �@�^���p�N�������ɂ́A���r�L�`���E�v���e�A�\�[���n�ƃI�[�g�t�@�W�[������܂��B �@�זE���łُ̈�ȃ^���p�N���̒~�ς�h������A�ߏ�Ƀ^���p�N�����������Ƃ���h�{�������������Ƃ��� �@�^���p�N���̃��T�C�N�����s������A�זE�����ɐN�������a����������r�����邱�ƂŐ��̂̍P�퐫�ێ��� �@�֗^���Ă��܂��B���̂ق��A�̔����̉ߒ��ł̃v���O�����זE����A�n���`���g���a�Ȃǂ̎����̔����A �@�זE�̂��}���ɂ��֗^���邱�Ƃ��m���Ă��܂��B �@�I�[�g�t�@�W�[�ł́A��x�ɑ����̃^���p�N�������������̂ŁA�o���N�����Ƃ��Ă�Ă��܂��B �@�I�[�g�t�@�W�[�̃��J�j�Y���̈Ⴂ���� �@�@(1)�}�N���I�[�g�t�@�W�[�A �@�@(2)�~�N���I�[�g�t�@�W�[�A �@�@(3)�V���y������ݐ��I�[�g�t�@�W�[��3�ɕ������Ă��܂��B 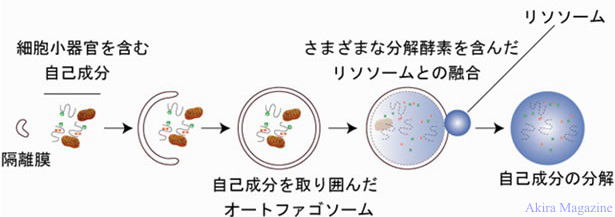 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�I�[�g�t�@�W�[�ɂ��`���������o�H�} �}�N���I�[�g�t�@�W�[�F �@�זE�������̃X�g���X�i�A�~�m�_�Q��̏�ԁA�ُ�^���p�N���̒~�ς�A�ہj�ɎN�����ƁA�זE������ �@�ꕔ�ŁA�ߏ�ɍ��ꂽ�^���p�N����ُ�^���p�N���Ƌ��Ƀ����������W�܂�A�I�[�g�t�@�S�\�[�� �@(Autophagosome�A�܂��̓I�[�g�t�@�W�[���E Autophagic vesicle)�ƌĂ��זE���\���i���Ɏ�荞�ށj�� �@�`�����͂��܂�B �@�I�[�g�t�@�S�\�[���ƍזE���̃��\�\�[�������Z�����N�����A�������ă��\�\�[���ƗZ���������̂� �@�I�[�g���\�\�[���ƌĂсA�I�[�g���\�\�[���̓����ŁA�I�[�g�t�@�S�\�[���ɗR�����镪�����ׂ��^���p�N���� �@�A���\�\�[���ɗR�����邳�܂��܂ȃ^���p�N�����y�f���������A���̌��ʁA�I�[�g�t�@�S�\�[���Ɏ�荞�܂�� �@�����^���p�N���̓A�~�m�_��y�v�`�h�ɕ��������B �~�N���I�[�g�t�@�W�[�F �@�ُ�^���p�N���ځA�t�E��\�\�[���Ɏ�荞�݁A���̓����ŕ�������@�\�B �V���y������ݐ��I�[�g�t�@�W�[�F �@�V���y�������ُ�^���p�N���Ɍ������邱�Ƃɂ���āA�t�E��\�\�[���ւ̎�荞�݂��s���A���̓����ŕ��� �@����@�\�B |
���r�L�`��
���r�L�`���E�v���e�A�\�[���@�@�i �`���������̈�� �j
| ���r�L�`�� (Ubiquitin)�́A�A�~�m�_����Ȃ�^���p�N���ŁA���̃^���p�N���̏C���ɗp�����A �@�ُ�ȃ^���p�N�����זE���番���A�����ADNA�C���A�|�߁A�V�O�i���`�B�Ȃǂ��܂��܂Ȑ������ۂ� �@�ւ��d�v�Ȗ����������Ă��܂��B �@���r�L�`���ɂ��W�����ꂽ�^���p�N�����v���e�A�\�[���ŕ��������r�L�`���E�v���e�A�\�[�� �V�X�e���� �@�Ă�Ă��܂��B �v���e�A�\�[�� (proteasome)�́A�^���p�N���̕������s������ȍy�f�����̂ł��B �@�זE��������A�Ɖu�����A�V�O�i���`�B�Ƃ������זE���̗l�X�ȓ����Ɋւ��@�\�ł��B ���r�L�`���E�v���e�A�\�[�� �V�X�e���̗���́A �@�@ �W�I�^���p�N�������r�L�`���ɂ��W�������B �@�A �W�����ꂽ�^���p�N���Ɍ����B �@�B �W�����ꂽ�^���p�N�����烆�r�L�`����藣���B�藣���ꂽ���r�L�`���͍ė��p����܂��B �@�C �W�����ꂽ�^���p�N���̗��̍\���������i�A���t�H�[���f�B���O�j�v���e�A�\�[�����ɑ��荞�܂�܂��B�@ �@�D �v���e�A�[�[�����ɂ��W�����ꂽ�^���p�N���͕�������܂��B 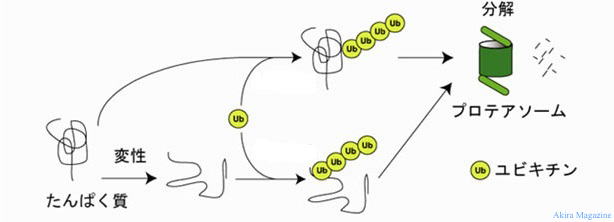 �@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@���r�L�`���ɂ��`���������o�H�} |
�G���h�T�C�g�|�V�X
�G���h�T�C�g�[�V�X �@( �����̎�荞�� �j
| �G���h�T�C�g�[�V�X�iEndocytosis)�Ƃ́A�זE������������Ɏ�荞�ލ�p�ł��B��荞�܂�镨�����̂� �@�ꍇ�ɂ͐H��p�ƌĂ�A�t�̂̏ꍇ�͈���p�Ƌ敪����Ă��܂��B�זE�O�a���́A�ٕ��A�A�|�g�[�V�X�זE �@�Ȃǂ��t�@�S�\�[���i�זE���ŕ����̕������s�����߂̏��튯�jphaosome�Ɏ�荞�݁A�����G���h�\�[���A �@����G���h�\�[�����o�R���ă��\�\�[���֗A������܂��B �@�t�@�S�T�C�g�[�V�X(phagocytosis)�@�F�������̂̎�荞�݂��H��p�̎��B �@�s�m�T�C�g�\�V�X�ipinocytosis�j�F�������t�̂̎�荞�݂�����p�̎��B |
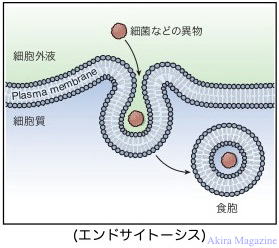 |
�G�L�\�T�C�g�|�V�X
�G�L�\�T�C�g�|�V�X�@�i �����̕��o �j
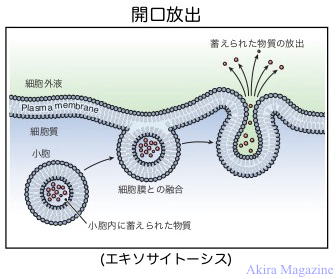
| �G�L�\�T�C�g�|�V�X�iExocytosis�j�Ƃ́A�זE����Z�������A���E���ɒ~����ꂽ�������זE������זE�O�� �@���o���邱�Ƃ������܂��B�Ⴆ��ƒ����_�o�n�ȂǂŐ_�o�`�B�����̕��o��A�����̓�����זE����� �@�z�������̕���͊J�����o�ɂ�肨���Ȃ���B |
 |











