難病 パーキンソン病患者とともに生きる。
Akira Magazinパーキンソン病 介護・症状日誌
神経細胞の数
大脳皮質の神経細胞の数はおよそ100億から180億くらいあり、一般には平均値をとって大脳皮質の神経細胞の数140億個であるとされています。
大脳皮質の神経細胞数は、チンパンジーで約80億、アカゲザル(ニホンザルの仲間)で約50億とされています。
注意すべき点は、大脳皮質の神経細胞の数であり、脳あるいは中枢神経(脳と脊髄)全体の数ではないということです。大脳の内側には神経細胞の集まった神経核があり、小脳や脊髄にも沢山の神経細胞があり,小脳だけでも1,000億以上の神経細胞があるという概算もあります。従って、中枢神経全体の神経細胞の数は1,000億と2,000億の間と推定されます。
身体の成長と共に神経細胞のネットワークも同時に成長。成長期間は、10代後半までと考えられています。
成長が止まり始めるのは4歳位からで、視覚、聴覚、本性感覚、次いで運動、言語に関する部位(運動野・言語野)、判断・計画を司る部位(前頭前野)の順序です。
最近の研究で、運動、反復、継続する事で新しい神経細胞ネットワークの活性化は可能と解ってきました。
細胞は1年位で新しい細胞と入れ替わりますが、脳、神経細胞、心臓(心筋細胞)は入れ替わり、再生できないと言われています。
Page Contents
| ①神経細胞 | ②シナプス |
| ③神経細胞の種類 | ④グリア細胞 |
| ⑤細胞変性 | ⑥ 神経細胞小器官 |
| ⑦エンドソーム | ⑧ミトコンドリア |
| ⑨オートファジー | ⑩ユビキチン、 プロテアソーム |
| ⑪エンドサイト-シス | ⑫エキソサイト-シス |
①神経細胞
*情報伝達ニューロンで、軸索・樹状突起に枝分かれ。
神経細胞は、情報伝達に特化した細胞です。一つの神経細胞からは長い「軸索」(じくさく、神経伝達物質の 投射)と、複雑に枝分かれしている「樹状突起」(じゅじょうとっき、神経伝達物質の受容)と呼ばれる突起が出ていて、これらの突起は別の神経細胞とつながり合い、複雑な神経回路網(ネットワーク)を形成しています。1つの神経細胞には約1万の樹状突起が付いていると言われています。また、細胞膜に包まれ核、ミトコンドリア、リソソームなどの細胞小器官をもっています。
軸索は、情報を送り出す突起で1本、軸索側枝と末端は枝分かれ(終末側枝)ています。
樹状突起は、情報を受け取る突起で細胞体から複数枝分かれしています。
情報の受取が少ない樹状突起は除去され、残った樹状突起の枝分かれを入力の多い方向に増やすことで、より効率的な神経回路が形成されます。
1個の神経細胞はそれぞれ1万個もの神経細胞と連絡を取り合っています。神経細胞内では、電気の流れが情報を伝え、神経細胞と神経細胞の接合部分はシナプス間隙と呼ばれるわずかな隙間があり、この部分では神経伝達物質が次の神経細胞に情報を伝達します。
グリア細胞は、神経系を構成する細胞では有りませんが、神経細胞に対して栄養素の供給や位置の固定を行っており、生体の内部や外部の環境因子の変化にかかわらず生体の状態が一定に保たれるという性質(恒常性)を持っています。
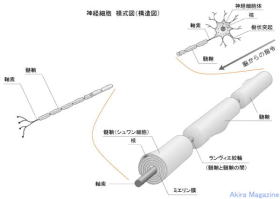
髄鞘化(ミエリン化)、神経伝達回路の形成に付いて:
神経細胞から次の神経細胞への神経伝達は電位で行われ、神経終末部分でシナプス小胞によりシナプス間隙にシナプス小胞の中の神経伝達物質が放出され樹状突起(神経伝達を受ける神経細胞)の受容体をへて神経伝達が行われる事が知られている。
神経細胞が新しく作られる時期は、20歳位とされていて神経伝達回路の形成は終わり、その後、使用頻度の少ない神経伝達回路(神経細胞)は消滅するとされていました。
最近の研究で、脳内の一部部位(海馬)では、神経伝達回路(神経細胞)の新たな形成が行えること、髄鞘化(ミエリン化)を促進させる事が研究により解ってきました。
髄鞘とは、神経細胞の軸索を包む円筒状の層(リン脂肪が巻きついた絶縁体)で軸索、髄鞘の径を太くする事(軸索成熟)により神経伝達をより高速化させる。
髄鞘化(ミエリン化)は、中枢神経ではグリア細胞(オリゴデンドロサイト)、末梢神経ではシュワン細胞から供給を受け軸索が一定の径に達すれば髄鞘形成が促進されます。
神経細胞は、情報伝達に特化した細胞です。一つの神経細胞からは長い「軸索」(じくさく、神経伝達物質の 投射)と、複雑に枝分かれしている「樹状突起」(じゅじょうとっき、神経伝達物質の受容)と呼ばれる突起が出ていて、これらの突起は別の神経細胞とつながり合い、複雑な神経回路網(ネットワーク)を形成しています。1つの神経細胞には約1万の樹状突起が付いていると言われています。また、細胞膜に包まれ核、ミトコンドリア、リソソームなどの細胞小器官をもっています。
軸索は、情報を送り出す突起で1本、軸索側枝と末端は枝分かれ(終末側枝)ています。
樹状突起は、情報を受け取る突起で細胞体から複数枝分かれしています。
情報の受取が少ない樹状突起は除去され、残った樹状突起の枝分かれを入力の多い方向に増やすことで、より効率的な神経回路が形成されます。
1個の神経細胞はそれぞれ1万個もの神経細胞と連絡を取り合っています。神経細胞内では、電気の流れが情報を伝え、神経細胞と神経細胞の接合部分はシナプス間隙と呼ばれるわずかな隙間があり、この部分では神経伝達物質が次の神経細胞に情報を伝達します。
グリア細胞は、神経系を構成する細胞では有りませんが、神経細胞に対して栄養素の供給や位置の固定を行っており、生体の内部や外部の環境因子の変化にかかわらず生体の状態が一定に保たれるという性質(恒常性)を持っています。
髄鞘化(ミエリン化)、神経伝達回路の形成に付いて:
神経細胞から次の神経細胞への神経伝達は電位で行われ、神経終末部分でシナプス小胞によりシナプス間隙にシナプス小胞の中の神経伝達物質が放出され樹状突起(神経伝達を受ける神経細胞)の受容体をへて神経伝達が行われる事が知られている。
神経細胞が新しく作られる時期は、20歳位とされていて神経伝達回路の形成は終わり、その後、使用頻度の少ない神経伝達回路(神経細胞)は消滅するとされていました。
最近の研究で、脳内の一部部位(海馬)では、神経伝達回路(神経細胞)の新たな形成が行えること、髄鞘化(ミエリン化)を促進させる事が研究により解ってきました。
髄鞘とは、神経細胞の軸索を包む円筒状の層(リン脂肪が巻きついた絶縁体)で軸索、髄鞘の径を太くする事(軸索成熟)により神経伝達をより高速化させる。
髄鞘化(ミエリン化)は、中枢神経ではグリア細胞(オリゴデンドロサイト)、末梢神経ではシュワン細胞から供給を受け軸索が一定の径に達すれば髄鞘形成が促進されます。
②シナプス
*神経細胞のつなぎ目。
神経細胞間のつなぎ目に関わる構造をシナプスと呼ばれています。
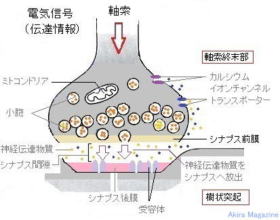
神経細胞の軸索より電位が伝わり、神経伝達物質が包まれたシナプス小胞が細胞膜(シナプス前膜)に接し膜電位依存性カルシウムイオンチャンネルが開きカルシウムが注入されシナプス小胞膜と軸索終末部の細胞膜とが融合してシナプス小胞内にある神経伝達物質がシナプス間隙に放出され、樹状突起の受容体に取り込まれ結合する事により情報伝達される。
シナプス小胞の大きさは50nmほど、シナプス間隙(かんげき)は20nmほど。(1nm=10億分の1m)、シナプス間隙の伝達時間は0.1~0.2ミリ秒ほど。
情報伝達機序:
1:神経細胞の軸索から電位が伝わり、軸索終末部に
到達する。
2:電位により軸索終末部に位置する電位依存性
カルシウム イオンチャネルが開く。
3:カルシウム イオンが軸索終末部に流入し、
シナプス小胞が細胞膜に接して神経伝達物質を細胞
外に開口 放出。
4:神経伝達物質はシナプス間隙を拡散し、樹状突起
(受側)の細胞膜上に分布する神経伝達物質受容体
に結合。
5:樹状突起側のシナプス後膜のイオン チャネルが
開き、細胞膜内外の電位差が変化します。
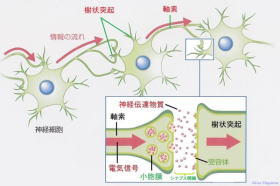
神経細胞 軸索・シナプス・樹状突起図
神経細胞間のつなぎ目に関わる構造をシナプスと呼ばれています。
神経細胞の軸索より電位が伝わり、神経伝達物質が包まれたシナプス小胞が細胞膜(シナプス前膜)に接し膜電位依存性カルシウムイオンチャンネルが開きカルシウムが注入されシナプス小胞膜と軸索終末部の細胞膜とが融合してシナプス小胞内にある神経伝達物質がシナプス間隙に放出され、樹状突起の受容体に取り込まれ結合する事により情報伝達される。
シナプス小胞の大きさは50nmほど、シナプス間隙(かんげき)は20nmほど。(1nm=10億分の1m)、シナプス間隙の伝達時間は0.1~0.2ミリ秒ほど。
情報伝達機序:
1:神経細胞の軸索から電位が伝わり、軸索終末部に
到達する。
2:電位により軸索終末部に位置する電位依存性
カルシウム イオンチャネルが開く。
3:カルシウム イオンが軸索終末部に流入し、
シナプス小胞が細胞膜に接して神経伝達物質を細胞
外に開口 放出。
4:神経伝達物質はシナプス間隙を拡散し、樹状突起
(受側)の細胞膜上に分布する神経伝達物質受容体
に結合。
5:樹状突起側のシナプス後膜のイオン チャネルが
開き、細胞膜内外の電位差が変化します。
神経細胞 軸索・シナプス・樹状突起図
③神経細胞の種類
神経細胞の働きを分類すると感覚、介在、運動の3種類に分けられます。簡単に言うと、外からの刺激を受け取るのが感覚神経細胞、刺激を筋肉に伝えるのが運動神経細胞、神経細胞 間での情報伝達をするのが介在神経細胞です。この介在神経細胞が、脳などの中枢神経系を形成しています。
情報伝達は電気信号(活動電位)により出力、入力と行われ、神経細胞(軸索、樹状突起)の末端では、シナプスと言う隙間がありその間は神経伝達物質で情報が伝達されています。
感覚神経細胞の種類
情報伝達は電気信号(活動電位)により出力、入力と行われ、神経細胞(軸索、樹状突起)の末端では、シナプスと言う隙間がありその間は神経伝達物質で情報が伝達されています。
感覚神経細胞の種類
| 種類 | 特徴 | はたらき |
| 感覚 神経細胞 |
受容器 (皮膚など感覚器) からの刺激を中枢に 伝える神経細胞。 細胞本体は背中側に 存在。 |
末梢から中枢 へ経路 を形成。 |
| 介在 神経細胞 |
神経細胞同しの 連絡 をする神経細胞で全体と しては短い。 脳、脊髄、交感神経節 の中枢に存在。 |
中枢神経系 を形成。 |
| 運動 神経細胞 |
中枢からの興奮を 効果器(筋肉や腺など) に伝える神経細胞。 |
中心から末梢 へ経路を形成 |
| グリア細胞 | 神経系の維持に関与する 細胞群で、神経細胞に 対し栄養素の供給や 神経細胞の位置固定 など、体温や血圧、 体液の浸透圧やpHなどを はじめ病原微生物や ウイルスといった異物の 排除、修復など生体機能 全般に及ぶ恒常性の維持 を担う細胞で、免疫系の ような振る舞いをする 細胞。 神経細胞数の約10倍と 推測されている。 |
視床下部の 指示により 自律神経系 や内分泌系 (ホルモン 分泌)を 担う |
④グリア細胞
中枢神経系や脊髄に存在して神経細胞(感覚・運動・介在)の維持に関与する細胞群で、神経細胞に対し、栄養素の供給や神経細胞の位置固定など、体温や血圧、体液の浸透圧やpHなどをはじめ病原微生物やウイルスといった異物の排除、修復など生体機能全般に及ぶ恒常性の維持を担う細胞で、免疫系のような振る舞いをする細胞。神経細胞数の約10倍、体積では脳の半分を占めると推測されている。
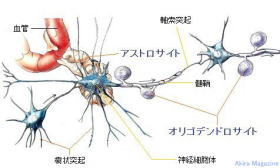
中枢神経系のグリア細胞図
グリア細胞の種類
中枢神経系のグリア細胞図
グリア細胞の種類
| 種類 (英語) |
種類 (日本語) |
特徴・はたらき | |
| ミクログリア microglia |
小膠細胞 しょうこう さいぼう |
中枢神経系で食作用 を示し免疫のほか 異常代謝物などの回収を担う細胞。 血管が損傷した時 以外は白血球が 入り込めないように ガードしている。 |
|
| アストロ サイト astrocyte |
星状膠細胞 せいじょうこう さいぼう |
中枢神経系に存在。 代謝産物を血液から 神経細胞まで輸送、 一方、間液から過剰 なカリウムイオン と神経伝達物質を 調整する。 組織学的に ・形質性星状膠細胞 ・線維性星状 ・膠細胞 に分類される。 神経幹細胞に由来。 |
|
| オリゴデンド ロサイト oligodendrocyte |
希突起膠細胞 きとっきこう さいぼう |
中枢神経系に存在。 軸索に巻きついて 髄鞘 (ずいしょう)を形成 および巻きついた 神経細胞の維持と 栄養補給の機能を 持つ。 |
|
| ependiomocyte, ependymal cell |
上衣細胞 じょういさいぼう |
中枢神経系に存在。 脳室系の壁を構成 する細胞である。 |
|
| シュワン細胞 Schwann cell |
鞘細胞 さやさいぼう |
末梢神経系に存在。 軸索に巻きついて 髄鞘を形成する。 1個のシュワン細胞 が1つの髄節を形成。 末梢神経が損傷した 時には再生の ため修復作用を 起こす。 |
|
| サテライト 細胞 satellite cell |
外套細胞 がいとう さいぼう |
筋組織に存在。 末梢神経系、 脊髄神経節や 交感神経節の 神経細胞体の周囲 を,外套のように取り 囲んで存在する細胞 ひとつ。神経細胞の 栄養補給機能を持つ。 |
|
⑤細胞変性
神経細胞の一部が傷つけられると、その場所よりも細胞体から遠い側は変性して壊れてしまう順行性変性。
細胞体のある側にも変性が進行することがあり、これを逆行性変性。また、神経細胞は互いに神経栄養因子などをやり取りしており、シナプスで接続している細胞が壊れた場合にも、神経栄養因子の不足から細胞死を起こすことがあります。この場合も、前シナプス細胞が死んだことにより後シナプス細胞が死ぬ場合を順行性変性、後シナプス細胞が死んだことにより前シナプス細胞が死ぬ場合を逆行性変性と呼ぶことがあります。
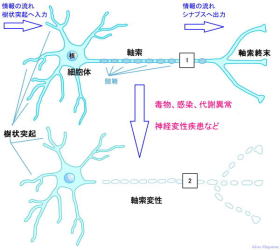
障害の程度が激しくて細胞体が死んでしまうと、その神経細胞はもはや再生不能。しかし、末梢神経(用語集参照)の場合、細胞体が生きていれば、再び軸索を伸ばして目的細胞との結合を回復できることが多く、その過程には、基底膜やシュワン細胞の関与が必要とされています。一方、末梢神経に比べて中枢神経はほとんど再生能力がなく、脳や脊髄の損傷は生涯に渡って後遺症を残すことが少なくない。
細胞体のある側にも変性が進行することがあり、これを逆行性変性。また、神経細胞は互いに神経栄養因子などをやり取りしており、シナプスで接続している細胞が壊れた場合にも、神経栄養因子の不足から細胞死を起こすことがあります。この場合も、前シナプス細胞が死んだことにより後シナプス細胞が死ぬ場合を順行性変性、後シナプス細胞が死んだことにより前シナプス細胞が死ぬ場合を逆行性変性と呼ぶことがあります。
障害の程度が激しくて細胞体が死んでしまうと、その神経細胞はもはや再生不能。しかし、末梢神経(用語集参照)の場合、細胞体が生きていれば、再び軸索を伸ばして目的細胞との結合を回復できることが多く、その過程には、基底膜やシュワン細胞の関与が必要とされています。一方、末梢神経に比べて中枢神経はほとんど再生能力がなく、脳や脊髄の損傷は生涯に渡って後遺症を残すことが少なくない。
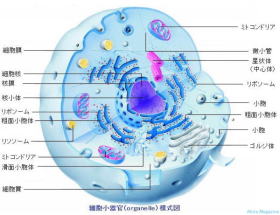
⑥ 神経細胞小器官
*核小体・小胞体・リボソーム・ゴジル体・リソソーム。
細胞小器官 (Organelle) は、 真核生物が持つ細胞小器官。核小体、小胞体、ゴルジ体、リソソーム、ミトコンドリア、ペルオキシソーム等の膜で囲まれた構造体。
小胞体、ゴルジ体、エンドソーム、リソソームは、小胞を介して細胞膜と連絡しあっている。
膜の内外で様々な物資の濃度をコントロールして、エネルギー生産(電子伝達系)、物質の貯蔵などを行っている。
・核小体(cell nucleus)には、遺伝情報であるDNAのほか、
核タンパク質、RNA(リボ核酸)が含まれています。
・リボソーム(Ribosome、タンパク質の製造工場)は、
細胞核のDNAより伝令RNA
(設計図、en:MessengerRNA)を読み取り、
転移RNA(en: Transfer RNA (TRNA)に結びついた
アミノ酸から所定のタンパク質に組み立てる。
(細胞膜を構成するタンパク質、分泌タンパク質を
合成)
・小胞体(endoplasmic reticulum)は、リボソームで
作られたタンパク質を立体構造(折りたたむ)に
関わっています。
・粗面小胞体(そめんしょうほうたい:
rough-surfaced endoplasmic reticulum, rER)では、
リボソームが付着しており、ゴルジ体やリソソーム、
小胞体、細胞膜等を構成するタンパク質および、
分泌タンパク質を合成。
・ゴジル体(Golgi body、タンパク質の最終検査工程)
は、リボソーム → 小胞体 → ゴジル体へ運ばれ、
分泌タンパク質や細胞外タンパク質の糖鎖修飾や、
小胞体(粗面小胞体)により生産された各種前駆体
タンパク質の化学的修飾(タンパク質を完成)を行い
タンパク質を分類し、分泌顆粒、リソソームあるいは
細胞膜にそれぞれ振り分ける働きをもつ。
・エンドソームは、物質の選別、分解再利用の機能。
別項に記載。
・ミトコンドリアに付いては別項に記載。
・リソソーム (Lysosome) は、 体高分子の分解と再
利用。また、細胞内の分解装置としてだけでなく、
他にも様々な機能を有しています。リソソームの
エキソサイト―シス(細胞外への物質の放出)は、
損傷した細胞膜の修復、細胞外マトリックスの分解な
どに関与。リソソーム膜の透過性亢進(lysosomal
membrane permeabilization)と細胞死との関連も
報告されています。また、
リソソームは細胞内の栄養状態(アミノ酸など)を
感知する場としても重要で、さらに細胞内のアミノ酸
濃度を感知するセンサータンパク質の多くリソソーム
膜上に局在しています。細胞内のアミノ酸濃度が上昇
すると、細胞成長・代謝・タンパク質合成などの
様々な細胞機能を制御する重要なシグナル因子で、
その活性化はリソソーム膜上で起こります。
リソソームの内腔は種々の加水分解酵素を含み、細胞
内外成分の分解機能を担っています。
分解基質はエンドサイトーシス、マクロオートファジー
などの経路によってリソソームに輸送されます。
分解された物体のうち有用なものは、細胞質に吸収
されます。不用物はエキソサイトーシス(細胞外へ
放出)によって廃棄されるか、残余小体(residual body)
として細胞内に留まります。
一次リソソーム(primary lysosome、一次水解小体):
分解基質を含まないリソソームを指し、内部が均一
な高電子密度顆粒。
二次リソソーム(secondary lysosome、二次水解
小体):
エンドソーム、ファゴソーム、オートファゴソームと
融合し分解基質を含んだ一次リソソーム。
二次リソソームは、さらに基質の輸送経路に従って
ファゴリソソーム(phagolysosome、食込融解小体)
オートリソソーム(autolysosome、自家食融解小体)と
呼ばれています。
細胞内には様々なリソソームがあり、その大きさ・
形状は様々に異なっています。
リソソームが含有する加水分解酵素群は酸性条件
下で効率良く働く性質を持っており、リソソーム内部
の水素イオン指数はプロトンポンプの働きによって
水素イオン指数値pH5 前後と酸性に保たれています。
(pH7が中性 )このことによって、中性状態の他の
細胞内区画ではリソソームが含む加水分解酵素は
不活性となり、不必要な反応を防いでいます。
ライソソーム(リリソーム)に障害が起ると
ミトコンドリアの鉄代謝に影響を及ぼし、糖脂質を
分解する
ライソソーム酵素、グルコセレブロシターゼ(GBA)の
欠乏によるゴーシェ病発病。
ゴーシェ病は、肝臓や脾臓の肥大、血小板減少に
により貧血症、骨が大変もろくなり、パーキンソン
ニズム
タンパク質の合成・異常タンパク質の処理、品質管理
タンパク質はアミノ酸が結合され作られます。新たに作ろうとするタンパク質は、その設計図(RNA)により作られます。RNAは細胞核のDNAによりコピーされリボソームで作られます。その後タンパク質は小胞体で折り畳み(立体構造)されますが、異常タンパク質、未完成タンパク質が作られた時に正常のタンパク質に戻すために分子シャペロンが働き正常な折り畳みに戻すのですが、大量に異常、未完成タンパク質が出来た場合(細胞体内、小胞体内)には、異常タンパク質の凝集により細胞自体が死にます。
この現象をフォールディング病(アルツハイマー型認知症、パーキンソン病、プリオン病等)です。この事を回避するために小胞体ストレス応答と言う機能があります。処理が行われる順番は、
① PERKにより異常タンパク質の合成停止、
② ATF6によりシャペロンRNAをリボソームに指示し
て小胞体に正常な折り畳みが指令される。
③ IRE1とXBP1によるタンパク質の分解と再生により
蛋白質の品質管理が行われています。
(転写因子:ATF6、XBP1は小胞体に存在して異常タンパク質が出来るとATF6が切り離され細胞核へ行き細胞 核がATF6を感知してシャペロンRNAをリボソームに指示して小胞体に正常な折り畳み指示が指令されます)
細胞小器官 (Organelle) は、 真核生物が持つ細胞小器官。核小体、小胞体、ゴルジ体、リソソーム、ミトコンドリア、ペルオキシソーム等の膜で囲まれた構造体。
小胞体、ゴルジ体、エンドソーム、リソソームは、小胞を介して細胞膜と連絡しあっている。
膜の内外で様々な物資の濃度をコントロールして、エネルギー生産(電子伝達系)、物質の貯蔵などを行っている。
・核小体(cell nucleus)には、遺伝情報であるDNAのほか、
核タンパク質、RNA(リボ核酸)が含まれています。
・リボソーム(Ribosome、タンパク質の製造工場)は、
細胞核のDNAより伝令RNA
(設計図、en:MessengerRNA)を読み取り、
転移RNA(en: Transfer RNA (TRNA)に結びついた
アミノ酸から所定のタンパク質に組み立てる。
(細胞膜を構成するタンパク質、分泌タンパク質を
合成)
・小胞体(endoplasmic reticulum)は、リボソームで
作られたタンパク質を立体構造(折りたたむ)に
関わっています。
・粗面小胞体(そめんしょうほうたい:
rough-surfaced endoplasmic reticulum, rER)では、
リボソームが付着しており、ゴルジ体やリソソーム、
小胞体、細胞膜等を構成するタンパク質および、
分泌タンパク質を合成。
・ゴジル体(Golgi body、タンパク質の最終検査工程)
は、リボソーム → 小胞体 → ゴジル体へ運ばれ、
分泌タンパク質や細胞外タンパク質の糖鎖修飾や、
小胞体(粗面小胞体)により生産された各種前駆体
タンパク質の化学的修飾(タンパク質を完成)を行い
タンパク質を分類し、分泌顆粒、リソソームあるいは
細胞膜にそれぞれ振り分ける働きをもつ。
・エンドソームは、物質の選別、分解再利用の機能。
別項に記載。
・ミトコンドリアに付いては別項に記載。
・リソソーム (Lysosome) は、 体高分子の分解と再
利用。また、細胞内の分解装置としてだけでなく、
他にも様々な機能を有しています。リソソームの
エキソサイト―シス(細胞外への物質の放出)は、
損傷した細胞膜の修復、細胞外マトリックスの分解な
どに関与。リソソーム膜の透過性亢進(lysosomal
membrane permeabilization)と細胞死との関連も
報告されています。また、
リソソームは細胞内の栄養状態(アミノ酸など)を
感知する場としても重要で、さらに細胞内のアミノ酸
濃度を感知するセンサータンパク質の多くリソソーム
膜上に局在しています。細胞内のアミノ酸濃度が上昇
すると、細胞成長・代謝・タンパク質合成などの
様々な細胞機能を制御する重要なシグナル因子で、
その活性化はリソソーム膜上で起こります。
リソソームの内腔は種々の加水分解酵素を含み、細胞
内外成分の分解機能を担っています。
分解基質はエンドサイトーシス、マクロオートファジー
などの経路によってリソソームに輸送されます。
分解された物体のうち有用なものは、細胞質に吸収
されます。不用物はエキソサイトーシス(細胞外へ
放出)によって廃棄されるか、残余小体(residual body)
として細胞内に留まります。
一次リソソーム(primary lysosome、一次水解小体):
分解基質を含まないリソソームを指し、内部が均一
な高電子密度顆粒。
二次リソソーム(secondary lysosome、二次水解
小体):
エンドソーム、ファゴソーム、オートファゴソームと
融合し分解基質を含んだ一次リソソーム。
二次リソソームは、さらに基質の輸送経路に従って
ファゴリソソーム(phagolysosome、食込融解小体)
オートリソソーム(autolysosome、自家食融解小体)と
呼ばれています。
細胞内には様々なリソソームがあり、その大きさ・
形状は様々に異なっています。
リソソームが含有する加水分解酵素群は酸性条件
下で効率良く働く性質を持っており、リソソーム内部
の水素イオン指数はプロトンポンプの働きによって
水素イオン指数値pH5 前後と酸性に保たれています。
(pH7が中性 )このことによって、中性状態の他の
細胞内区画ではリソソームが含む加水分解酵素は
不活性となり、不必要な反応を防いでいます。
ライソソーム(リリソーム)に障害が起ると
ミトコンドリアの鉄代謝に影響を及ぼし、糖脂質を
分解する
ライソソーム酵素、グルコセレブロシターゼ(GBA)の
欠乏によるゴーシェ病発病。
ゴーシェ病は、肝臓や脾臓の肥大、血小板減少に
により貧血症、骨が大変もろくなり、パーキンソン
ニズム
タンパク質の合成・異常タンパク質の処理、品質管理
タンパク質はアミノ酸が結合され作られます。新たに作ろうとするタンパク質は、その設計図(RNA)により作られます。RNAは細胞核のDNAによりコピーされリボソームで作られます。その後タンパク質は小胞体で折り畳み(立体構造)されますが、異常タンパク質、未完成タンパク質が作られた時に正常のタンパク質に戻すために分子シャペロンが働き正常な折り畳みに戻すのですが、大量に異常、未完成タンパク質が出来た場合(細胞体内、小胞体内)には、異常タンパク質の凝集により細胞自体が死にます。
この現象をフォールディング病(アルツハイマー型認知症、パーキンソン病、プリオン病等)です。この事を回避するために小胞体ストレス応答と言う機能があります。処理が行われる順番は、
① PERKにより異常タンパク質の合成停止、
② ATF6によりシャペロンRNAをリボソームに指示し
て小胞体に正常な折り畳みが指令される。
③ IRE1とXBP1によるタンパク質の分解と再生により
蛋白質の品質管理が行われています。
(転写因子:ATF6、XBP1は小胞体に存在して異常タンパク質が出来るとATF6が切り離され細胞核へ行き細胞 核がATF6を感知してシャペロンRNAをリボソームに指示して小胞体に正常な折り畳み指示が指令されます)
⑦エンドソーム
*細胞内の膜物質輸送 。
エンドソーム (Endosome)は、エンドサイトーシスによって細胞内へと取り込まれた様々な物質の選別・分解・再利用など
を制御。エンドソームは形態的な特徴や機能的などの特徴をもとに、
初期エンドソーム(early endosome)
後期エンドソーム(late endosome)
リサイクリングエンドソーム(recycling endosome)
に大別される。
エンドソームは膜輸送の中継地点として機能することで細胞内の物質輸送を司ることから、神経機能をはじめとする様々な生命現象において重要な役割を果たしている。
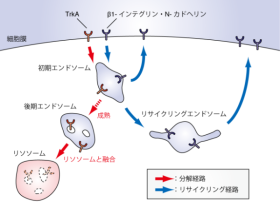
物質(TrkA)は分解経路へ、細胞膜で再利用される物質(β1-インテグリン、N-カドヘリン)はリサイクリング経路へと選別されていく。初期エンドソームにおいて分解経路へと選別された物質は、初期エンドソームに留まる。
そして初期エンドソームが後期エンドソームへと成熟すると、後期エンドソームがリソソームと融合し、内腔小胞ごと膜貫通型のタンパク質も分解される。
初期エンドソームにおいてリサイクリング経路へと選別された物質は、初期エンドソームから直接、あるいは核近傍のリサイクリングエンドソームへと輸送されたのち、細胞膜へと戻され再利用される。
エンドソーム (Endosome)は、エンドサイトーシスによって細胞内へと取り込まれた様々な物質の選別・分解・再利用など
を制御。エンドソームは形態的な特徴や機能的などの特徴をもとに、
初期エンドソーム(early endosome)
後期エンドソーム(late endosome)
リサイクリングエンドソーム(recycling endosome)
に大別される。
エンドソームは膜輸送の中継地点として機能することで細胞内の物質輸送を司ることから、神経機能をはじめとする様々な生命現象において重要な役割を果たしている。
物質(TrkA)は分解経路へ、細胞膜で再利用される物質(β1-インテグリン、N-カドヘリン)はリサイクリング経路へと選別されていく。初期エンドソームにおいて分解経路へと選別された物質は、初期エンドソームに留まる。
そして初期エンドソームが後期エンドソームへと成熟すると、後期エンドソームがリソソームと融合し、内腔小胞ごと膜貫通型のタンパク質も分解される。
初期エンドソームにおいてリサイクリング経路へと選別された物質は、初期エンドソームから直接、あるいは核近傍のリサイクリングエンドソームへと輸送されたのち、細胞膜へと戻され再利用される。
⑧ミトコンドリア
*エネルギーの産生、アポトーシス誘導に関与。
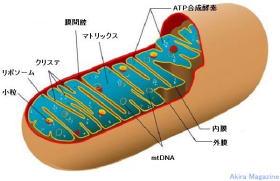
ミトコンドリア (mitochondria)は、真核生物の細胞小器官の一つ。二重の生体膜からなり、独自のDNA(mtDNA)を持ち、分裂、増殖する。ミトコンドリアは細胞の中で酸素(血液により運ばれた)により糖や脂肪を燃料する事により熱の産生(体温)、DNA損傷、ストレスによるアポトーシス誘導、調節、カルシウム貯蔵、細胞内のカルシウム濃度の調整に関与している。(ミトコンドリアDNA(mtDNA)はアデノシン三リン酸(ATP)以外のエネルギー産生に関与)mtDNAは、母系遺伝子で母親から受け継がれます。
ミトコンドリアの構造は、外膜と内膜の脂質膜に包まれており、内膜の内側はマトリックスと呼ばれ、内膜から平板状(クリステ)がある。外膜には、膜タンパク質(ポリン)があり、分子量5000以下の分子が透過できるようなチャネルを形成している。
内膜には、酸化的リン酸に関わる呼吸鎖複合体などの酵素群が規則的に配列している。また、マトリックスへのタンパク質輸送装置やミトコンドリアの分裂・融合に関わるタンパク質群などが存在。
マトリックスには、TCA回路(クレブス回路・クエン酸回路)やβ酸化などミトコンドリアの代謝機能に関わる酵素群、リボソーム、tRNA、転写因子や翻訳因子なども存在している。
ミトコンドリアの機能は、電子伝達系、リン酸化によるATPの産生(ADPのリン酸化)。細胞の活動に必要なエネルギーのほとんどは直接、間接的にミトコンドリアからATPの形で供給される。 その他の機能としては、ステロイドやヘムの合成などを含む様々な代謝、カルシウムや鉄の細胞内濃度の調節、細胞周期やアポトーシスの調節などにも大きく関わっている。
ミトコンドリア模式図
ミトコンドリア (mitochondria)は、真核生物の細胞小器官の一つ。二重の生体膜からなり、独自のDNA(mtDNA)を持ち、分裂、増殖する。ミトコンドリアは細胞の中で酸素(血液により運ばれた)により糖や脂肪を燃料する事により熱の産生(体温)、DNA損傷、ストレスによるアポトーシス誘導、調節、カルシウム貯蔵、細胞内のカルシウム濃度の調整に関与している。(ミトコンドリアDNA(mtDNA)はアデノシン三リン酸(ATP)以外のエネルギー産生に関与)mtDNAは、母系遺伝子で母親から受け継がれます。
ミトコンドリアの構造は、外膜と内膜の脂質膜に包まれており、内膜の内側はマトリックスと呼ばれ、内膜から平板状(クリステ)がある。外膜には、膜タンパク質(ポリン)があり、分子量5000以下の分子が透過できるようなチャネルを形成している。
内膜には、酸化的リン酸に関わる呼吸鎖複合体などの酵素群が規則的に配列している。また、マトリックスへのタンパク質輸送装置やミトコンドリアの分裂・融合に関わるタンパク質群などが存在。
マトリックスには、TCA回路(クレブス回路・クエン酸回路)やβ酸化などミトコンドリアの代謝機能に関わる酵素群、リボソーム、tRNA、転写因子や翻訳因子なども存在している。
ミトコンドリアの機能は、電子伝達系、リン酸化によるATPの産生(ADPのリン酸化)。細胞の活動に必要なエネルギーのほとんどは直接、間接的にミトコンドリアからATPの形で供給される。 その他の機能としては、ステロイドやヘムの合成などを含む様々な代謝、カルシウムや鉄の細胞内濃度の調節、細胞周期やアポトーシスの調節などにも大きく関わっている。
ミトコンドリア模式図
⑨オートファジー
*蛋白質分解の一つ。
オートファジー (Autophagy)は、細胞が持っている、細胞内のタンパク質を分解するための仕組みの一つで、タンパク質分解には、ユビキチン・プロテアソーム系とオートファジーがあります。
細胞内での異常なタンパク質の蓄積を防いだり、過剰にタンパク質合成したときや栄養環境が悪化したときにタンパク質のリサイクルを行ったり、細胞質内に侵入した病原微生物を排除することで生体の恒常性維持に関与しています。このほか、個体発生の過程でのプログラム細胞死や、ハンチントン病などの疾患の発生、細胞のがん化抑制にも関与することが知られています。
オートファジーでは、一度に多くのタンパク質が分解されるので、バルク分解とも呼ばれています。
オートファジーのメカニズムの違いから
(1)マクロオートファジー、
(2)ミクロオートファジー、
(3)シャペロン介在性オートファジーの3つに分けられ ています。
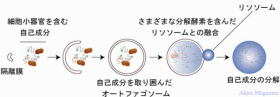
オートファジーによる蛋白質分解経路図
マクロオートファジー:
細胞がある種のストレス(アミノ酸飢餓の状態、異常タンパク質の蓄積や、細菌)に晒されると、細胞質中の一部で、過剰に作られたタンパク質や異常タンパク質と共にリン脂質が集まり、オートファゴソーム(Autophagosome、またはオートファジー小胞 Autophagic vesicle)と呼ばれる細胞内構造(膜に取り込む)の形成がはじまる。
オートファゴソームと細胞内のリソソームが膜融合を起こし、こうしてリソソームと融合したものをオートリソソームと呼び、オートリソソームの内部で、オートファゴソームに由来する分解すべきタンパク質と、リソソームに由来するさまざまなタンパク分解酵素が反応し、この結果、オートファゴソームに取り込まれていたタンパク質はアミノ酸やペプチドに分解される。
ミクロオートファジー:
異常タンパク質を直接、液胞やリソソームに取り込み、その内部で分解する機構。
シャペロン介在性オートファジー:
シャペロンが異常タンパク質に結合することによって、液胞やリソソームへの取り込みを行い、その内部で分解する機構。
オートファジー (Autophagy)は、細胞が持っている、細胞内のタンパク質を分解するための仕組みの一つで、タンパク質分解には、ユビキチン・プロテアソーム系とオートファジーがあります。
細胞内での異常なタンパク質の蓄積を防いだり、過剰にタンパク質合成したときや栄養環境が悪化したときにタンパク質のリサイクルを行ったり、細胞質内に侵入した病原微生物を排除することで生体の恒常性維持に関与しています。このほか、個体発生の過程でのプログラム細胞死や、ハンチントン病などの疾患の発生、細胞のがん化抑制にも関与することが知られています。
オートファジーでは、一度に多くのタンパク質が分解されるので、バルク分解とも呼ばれています。
オートファジーのメカニズムの違いから
(1)マクロオートファジー、
(2)ミクロオートファジー、
(3)シャペロン介在性オートファジーの3つに分けられ ています。
オートファジーによる蛋白質分解経路図
マクロオートファジー:
細胞がある種のストレス(アミノ酸飢餓の状態、異常タンパク質の蓄積や、細菌)に晒されると、細胞質中の一部で、過剰に作られたタンパク質や異常タンパク質と共にリン脂質が集まり、オートファゴソーム(Autophagosome、またはオートファジー小胞 Autophagic vesicle)と呼ばれる細胞内構造(膜に取り込む)の形成がはじまる。
オートファゴソームと細胞内のリソソームが膜融合を起こし、こうしてリソソームと融合したものをオートリソソームと呼び、オートリソソームの内部で、オートファゴソームに由来する分解すべきタンパク質と、リソソームに由来するさまざまなタンパク分解酵素が反応し、この結果、オートファゴソームに取り込まれていたタンパク質はアミノ酸やペプチドに分解される。
ミクロオートファジー:
異常タンパク質を直接、液胞やリソソームに取り込み、その内部で分解する機構。
シャペロン介在性オートファジー:
シャペロンが異常タンパク質に結合することによって、液胞やリソソームへの取り込みを行い、その内部で分解する機構。
⑩ユビキチン、プロテアソーム
*蛋白質分解の一つ。
ユビキチン (Ubiquitin)は、アミノ酸からなるタンパク質で、他のタンパク質の修飾に用いられ、異常なタンパク質を細胞から分解、除去、DNA修復、翻訳調節、シグナル伝達などさまざまな生命現象に関わる重要な役割を持っています。
ユビキチンにより標識されたタンパク質をプロテアソームで分解をユビキチン・プロテアソーム システムと呼ばれています。
プロテアソーム (proteasome)は、タンパク質の分解を行う巨大な酵素複合体です。
細胞周期制御、免疫応答、シグナル伝達といった細胞中の様々な働きに関わる機構です。
ユビキチン・プロテアソーム システムの流れは、
① 標的タンパク質がユビキチンにより標識される。
② 標識されたタンパク質に結合。
③ 標識されたタンパク質からユビキチンを切り離す。
切り離されたユビキチンは再利用されます。
④ 標識されたタンパク質の立体構造を解き(アンフォール
ディング)プロテアソーム内に送り込まれます。
⑤ プロテアーゼ活性により標識されたタンパク質は分解
されます。
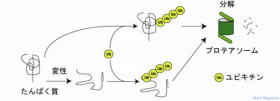
ユビキチンによる蛋白質分解経路図
ユビキチン (Ubiquitin)は、アミノ酸からなるタンパク質で、他のタンパク質の修飾に用いられ、異常なタンパク質を細胞から分解、除去、DNA修復、翻訳調節、シグナル伝達などさまざまな生命現象に関わる重要な役割を持っています。
ユビキチンにより標識されたタンパク質をプロテアソームで分解をユビキチン・プロテアソーム システムと呼ばれています。
プロテアソーム (proteasome)は、タンパク質の分解を行う巨大な酵素複合体です。
細胞周期制御、免疫応答、シグナル伝達といった細胞中の様々な働きに関わる機構です。
ユビキチン・プロテアソーム システムの流れは、
① 標的タンパク質がユビキチンにより標識される。
② 標識されたタンパク質に結合。
③ 標識されたタンパク質からユビキチンを切り離す。
切り離されたユビキチンは再利用されます。
④ 標識されたタンパク質の立体構造を解き(アンフォール
ディング)プロテアソーム内に送り込まれます。
⑤ プロテアーゼ活性により標識されたタンパク質は分解
されます。
ユビキチンによる蛋白質分解経路図
⑪エンドサイト-シス
*物質の取り込み。
エンドサイトーシス(Endocytosis)とは、細胞が物質を内部に取り込む作用です。取り込まれる物質が個体の場合には食作用と呼ばれ、液体の場合は飲作用と区分されています。細胞外病原体、異物、アポトーシス細胞などをファゴソーム(細胞内で物質の分解を行うための小器官)phaosomeに取り込み、初期エンドソーム、後期エンドソームを経由してリソソームへ輸送されます。
ファゴサイトーシス(phagocytosis) :物質が個体の取り込みの食作用の事。
ピノサイト―シス(pinocytosis):物質が液体の取り込みの飲作用の事。
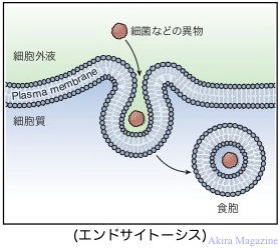
エンドサイトーシス(Endocytosis)とは、細胞が物質を内部に取り込む作用です。取り込まれる物質が個体の場合には食作用と呼ばれ、液体の場合は飲作用と区分されています。細胞外病原体、異物、アポトーシス細胞などをファゴソーム(細胞内で物質の分解を行うための小器官)phaosomeに取り込み、初期エンドソーム、後期エンドソームを経由してリソソームへ輸送されます。
ファゴサイトーシス(phagocytosis) :物質が個体の取り込みの食作用の事。
ピノサイト―シス(pinocytosis):物質が液体の取り込みの飲作用の事。
⑫エキソサイト-シス
* 物質の放出 。
エキソサイト-シス(Exocytosis)とは、細胞膜を融合させ、小胞内に蓄えられた物質を細胞内から細胞外に放出することを言います。例えると中枢神経系などで神経伝達物質の放出や、多くの内分泌細胞からのホルモンの分泌は開口放出によりおこなわれる。
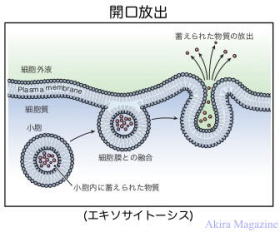
エキソサイト-シス(Exocytosis)とは、細胞膜を融合させ、小胞内に蓄えられた物質を細胞内から細胞外に放出することを言います。例えると中枢神経系などで神経伝達物質の放出や、多くの内分泌細胞からのホルモンの分泌は開口放出によりおこなわれる。
Akira Magazine
〒612-8065
Fhimi-ku Kyoto ctiy