中枢神経系のおはなしCentral nerve system
中枢神経の役割・機能
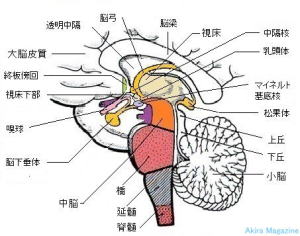
神経系には中枢神経系と末梢神経系があります。
目や耳、手足、体幹、内臓などから情報を送る末梢神経。末梢神経から情報を受け取り、感覚、運動、意思、情緒、反射、呼吸など、コントロールする中枢神経。
多数の神経細胞が集まっている領域です。
(モノアミン系 + コリン系)
Page Contents (詳細は、下線部分をクリック)
・アセチルコリン系 ・アドレナリン系
・ セロトニン系 ・ドーパミン系
・ノルアドレナリン系 ・ヒスタミン系
中枢神経系に関わる脳部位と神経伝達物質
| 脳の名称 | 脳部位 | モノアミンの神経伝達物質 | |||
| 脳 | 大脳 | 大脳皮質 | ・前頭葉・頭頂葉 ・側頭葉・後頭葉 |
アセチルコリン系・(GABA) | |
| 大脳基底核 | ・線条体・淡蒼球 ・視床下核・黒質 |
ー | |||
| 大脳辺縁系 | ・帯状回・扁桃体 ・海馬・側坐核 |
アドレナリン系・(Ad) | |||
| 前脳基底部 | ・ マイネルト基底核 | セロトニン系・(5HT) | |||
| 髄質 | ・白質・副腎髄質 | ドーパミン系・(DA) | |||
| 間脳 | ・視床 ・視床下部 | ノルアドレナリン系・(NA) | |||
| 脳幹 | ・中脳・赤核・腹側被蓋野 ・橋・延髄 |
ヒスタミン・(H) | |||
| 小脳 | ー | ー | |||
| 脊髄 | ー | ー | ー | ||
アセチルコリン神経系
アセチルコリン神経系 (Acetylcholine nerve)
| アセチルコリンの合成 アセチルコリン(ACh)は、コリンとアセチルから、コリンアセチルトランスフェラーゼ(ChAT)の作用で作ら れる。 アセチルコリンは、コリンエステラーゼ(AchE)の作用で、コリンと酢酸に分解される。自己受容体を 持たないが蓄積性はない。 アセチルコリンの作用 アセチルコリンは、運動神経の神経終末、交感神経と副交感神経の神経節、副交感神経(迷走神経)の神経 終末、交感神経(汗腺のみ)の神経終末における伝達物質として機能しています。 アセチルコリン受容体 アセチルコリン受容体は、ムスカリン受容体(代謝調節型)、ニコチン受容体(イオンチャネル型)の二つ に大別され、アセチルコリンによって刺激されるコリン作動性受容体とも呼ばれる。末梢では副交感神経の 神経終末に存在し、副交感神経の活動を制御。 ・ニコチン受容体は1種類。 ・ムスカリン受容体は5種類。 M1: 脳(皮質、海馬)、腺、交感神経に分布。 M2: 心臓、後脳、平滑筋に分布。 M3: 平滑筋、腺、脳に分布。 M4: 脳(前脳、線条体)に分布。 M5: 脳(黒質)、眼に分布。 M2と結合で、洞房結節に作用し、心臓の心拍数を低下させる。 M3結合で、血管平滑筋では拡張し、血圧が低下する。 消化器では、消化液(胃酸・唾液)の分泌を促進する。 気管支平滑筋は収縮する。 眼では、縮瞳し、眼圧が低下する。 膀胱は収縮し排尿を促す。 膀胱にはM2/M3受容体が多い。 アセチルコリンがM2と結合すれば、細胞機能が 抑制され、M3と結合すれば促進される アセチルコリンの神経核
アセチルコリンの介在神経 線条体、側坐核、嗅結節、カレハ核に小型の介在神経が存在する。 |
アドレナリン神経系
アドレナリン神経系 (Adrenaline nerve)
| アドレナリンの合成 アドレナリンは副腎髄質にてノルアドレナリンからN-メチルトランスフェラーゼによって合成される。 外部ストレスにより刺激を受けた視床下部室傍核ニューロンが交感神経を賦活化し、交感神経終末から ノルアドレナリンの分泌を促進する。 交感神経から分泌されたノルアドレナリンが副腎髄質からのアドレナリン分泌を促す。 (副腎髄質からはAd:NAd=8:2程度の割合で分泌される) 神経核 ノルアドレナリン神経として副腎へ投射して、ノルアドレナリンの刺激で副腎からのアドレナリンが分泌 される。 アドレナリンを分泌する副腎は腎臓の隣にあるため、中枢には存在していない。一方、ノルアドレナリンを 分泌する青斑核を中心とした神経核は全て中枢に存在。 アドレナリンの作用 ノルアドレナリンとアドレナリンに共通する作用としては、交感神経の受容体であるα、β受容体を刺激して 血管を収縮させたり、心悸亢進させたりすること。 アドレナリンは、β2を刺激を介してグリコーゲン分解 → 血糖上昇作用がある。 ノルアドレナリンは、αやβ受容体を介して血管収縮や気管支収縮といった作用を示すだけでなく、脳内で セロトニンを抑制する縫線核へ投射したり、マイネルト基底核に投射してアセチルコリンによる大脳皮質の 賦活作用を増強したりと神経伝達物質として機能する。 アドレナリン受容体 アドレナリン受容体の種類は、α1、α2、βの三種類に分類されている。
アドレナリンの神経核
|
||||||||||||||||||||
セロトニン神経系
セロトニン神経系 (Serotonin nerve)
| セロトニンの産生・排出 トリプトファンからトリプトファン水酸化酵素により、5-水酸化トリプトファン(5-HTP)へ、 5-水酸化トリプトファン脱炭酸酵素によりセロトニンを産生。 神経終末から分泌されたセロトニンは、神経終末のセロトニントランスポーターから再吸収され、残り 約90%以上は,モノアミン酸化酵素A(MAO-A)により速やかに分解されて、5-水酸化インドール酢酸 (5-HIAA))へと代謝され、尿から排泄される。 脳内で水酸化酵素を持つのは、縫線核群の5HT神経か松果体のメラトニン産生細胞だけ。 セロトニンの分布 消化管粘膜(腸クロム親和性細胞)に約90%、血小板に約8%、中枢神経系に約2%。 セロトニンの分泌・抑制要因 セロトニンの分泌亢進因子は、歩行、咀嚼、呼吸のリズム運動と太陽光である。 光は網膜から縫線核へ直接の軸索投射で、5HT神経を賦活する。 抑制要因は、慢性ストレス。ストレスによって活性化される神経である室傍核CRH神経が縫線核に投射し ていて、5-HT神経の活動を抑制する。(一過性の覚醒・ストレス刺激に対しては影響されない) セロトニンの作用(セロトニン受容体は、14種類あり、作用は多種多様)
セロトニン受容体 5-HT3受容体のみが、イオンチャンネル共役型受容体で、残りはGタンパク質結合受容体です。
5-HT1Aでセロトニンが減少すると衝動的で暴力的でまた、 憂鬱(ゆううつ)となり、自制心がなくなる。 5-HT2Aにセロトニンが結合するとウツ病となる。 5-HT2Cにセロトニンが結合すると満腹感が得られる。 抗ウツ剤でセロトニンに作用するものは、5-HT2Aと2Cのセロトニン量を減らすため、ウツは改善されるが、 いくら食べても満腹感が得られぜず太ってしまう。 セロトニンの神経核
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
ドーパミン神経系
ドーパミン神経系 (Dopamin nerve)
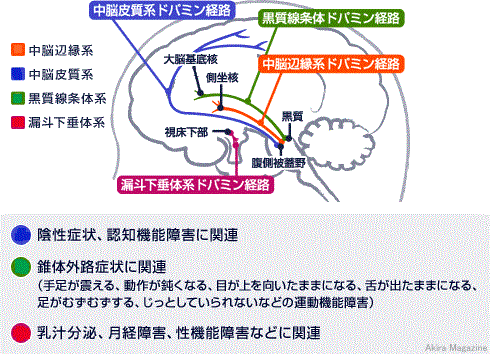
| ドーパミンの合成 チロシン(アミノ酸)からチロシン水酸化酵素によってレボトパ(L-Dopa)、ドーパ脱炭酸酵素によって ドーパミンに合成される。 ドーパミンは、酸化還元酵素によりノルアドレナリンへ合成され、基転移酵素によりアドレナリンに合成 される。 また、ノルアドレナリンは、カテコール-O-メチル転移酵素(COMT)によってノルメタネフリンへ代謝され、 モノアミン酸化酵素(MAO)によって、3-メトキシ-4水酸化フェニールグリコール(MHPG)へと分解され代謝 される。 ドーパミンの神経核 (黒質・緻密部 → 線条体経路と腹側被蓋野 → 辺縁系、皮質経路が重要)
ドーパミン受容体 ド-パミン受容体は、D1、D2、D3、D4、D5受容体と5種類あるとわかっています。 ドーパミンが受容体と結合すると神経細胞に電位変化が起こったり、細胞内の情報伝達系が動いてさまざまな 変化を起こします。受容体は、興奮性、抑制性と2つに分類されていますが、まだ不明点が多く単純では なさそうです。 興奮性を担う受容体:D1、D5受容体。抑制性を担う受容体:D2、D3、D4受容体。
|
|||||||||||||||||||||||||||||||||||||
ノルアドレナリン神経系
ノルアドレナリン神経系 (Noradrenaline nerve)
| ノルアドレナリンの合成 ドーパミンより酸化還元酵素によりノルアドレナリンへ合成される。 伝達 外側被蓋NAd神経系は中心被蓋束 (腹側NAd束) を介して上行し、視床、視床下部、前脳基底部 (主としてマイネルト基底核)などへ投射する。 青斑核NAd神経は、背側被蓋 (NAd) 束を上行し、内側前脳束を介して、視床、視床下部、中隔、海馬、 扁桃体などに軸索側枝を投射。 また、大脳皮質の全域に神経終末を投射され、さらに小脳や脊髄にも投射する。 ノルアドレナリンの作用 ストレス反応により活性化され、視床下部室傍核を中心としコルチゾールの分泌し交感神経の活性化が 起こる。 ・前脳基底部へ投射し、アセチルコリン分泌を介して覚醒反応に関わる。 ・扁桃体への投射は不安や注意など情動的情報を記憶・付加し、 情動回路として前頭前野へ投射、不安 ・恐怖・パニック等の行動を引き起こす。 ノルアドレナリンの神経核
|
ヒスタミン神経系
ヒスタミン神経系 (Histamine)
| ヒスタミンの合成 ヒスタミンは、ヒスチジンからヒスチジン脱炭酸酵素により生合成される。 ヒスチジン脱炭酸酵素はピリドキサールリン酸を補酵素として必要とする。ほ乳類脳においてヒスタミンの 多くはヒスタミンメチル基転移酵素によりメチル化され、さらにモノアミン酸化酵素B(MAO-B)により 酸化され、メチルイミダゾール酢酸となり排出される。(肥満細胞などで産生) 伝達 ヒスタミン作動性神経細胞は、視床下部の結節乳頭体核に存在。投射先は脳全域に及ぶが、その密度は低い。 ヒスタミンの作用 末梢神経系では胃酸分泌、免疫反応、子宮や気管支などの不随意筋を収縮。 中枢神経系においては神経伝達物質としても働く。脳におけるヒスタミンの作用は、覚醒、興奮などをもた らし、血圧や痛みの調節に関与。 ヒスタミン受容体
ヒスタミンの神経核
|











